

Neogene spore-pollen assemblages and paleovegetation succession of Borehole ZKA02 in Beihai, Guangxi
-
摘要:
通过对广西壮族自治区北海市新生代钻孔ZKA02中的孢粉化石的系统研究,初步恢复了该地区新近纪植被变化历史和气候变化过程。根据孢粉化石数据,将该钻孔自下而上划分出4个孢粉组合。早中新世—中中新世该地区的植被以落叶阔叶混交林−湿生草甸植被为主,气候偏温和半湿润;中中新世末期—晚中新世早期以针叶阔叶混交林−湿生草甸植被、温和半湿润气候为主;晚中新世中期为落叶阔叶混交林−湿生草甸植被,针叶树和蕨类植物逐渐增多,整体气候为温暖湿润型;晚中新世晚期—上新世为落叶阔叶混交林−湿生草甸植被,热带雨林和亚热带低山常绿阔叶树种明显增加,蕨类分子明显减少,相较于前三阶段,该阶段气候更加干旱,整体气候为偏温暖半干旱型。孢粉组合所反映的新近纪气候变化特征与全球气候变化趋势具有较好的可比性。
Abstract:The history of vegetation and climate change during the Cenozoic has been reconstructed by systematically analyzing the sporopollen assemblage from Borehole ZKA02 in Beihai, Guangxi. Through the analysis of spore-pollen data, four sporopollen assemblages can be recognized. From the Early Miocene to the Middle Miocene, this area was covered mainly by deciduous broad-leaved mixed forest–wet meadow vegetation, with a mild and semi-humid climate. From the end of the Middle Miocene to the early Late Miocene, the study area was dominated by coniferous and broad-leaved mixed forest-wet meadow vegetation, with mild and semi-humid. The middle Late Miocene was a mixed deciduous broad-leaved forest–wet meadow vegetation, with increasing conifers and ferns, and the climate was warm and humid. In the late Late Miocene to Pliocene, it was deciduous broad-leaved mixed forest–wet meadow vegetation. The species of evergreen broad-leaved trees in tropical rain forests and subtropical low mountains increased obviously, and the fern molecules decreased obviously. The climate in the Late Pliocene was warm and semi-arid, drier than the previous three stages. Spore-pollen assemblages reflected the climate change characteristics during the Neogene, which have good comparability with the trend of global climate change.
-
Key words:
- spore-pollen assemblage /
- Neogene /
- paleo-vegetation /
- paleoclimate /
- Beihai
-
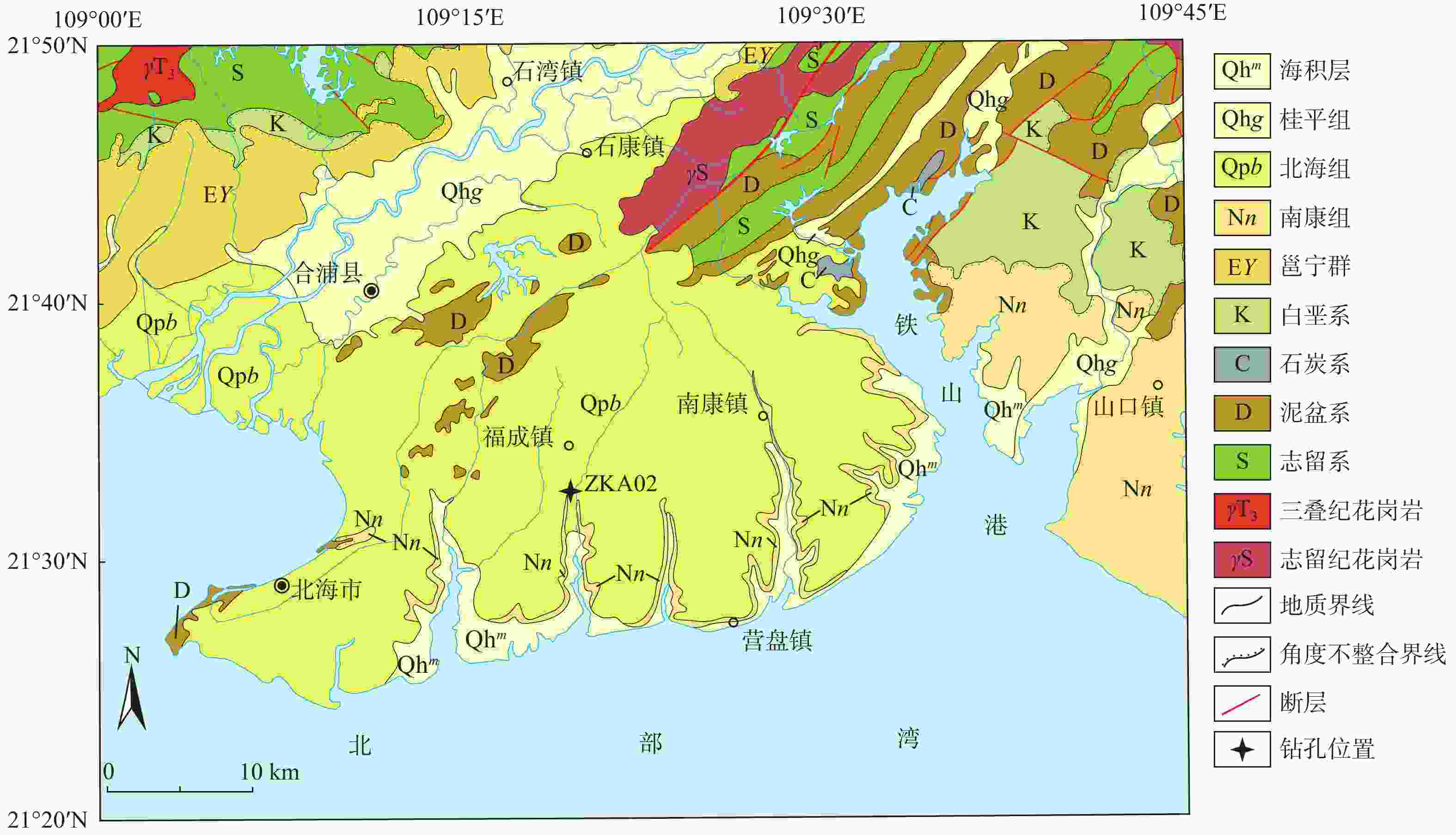
图 1 研究区地质简图(据广西壮族自治区地质调查研究院,2017修改)
Figure 1. Geologic sketch of the study area (modified from Guangxi Institute of Geological Survey, 2017)
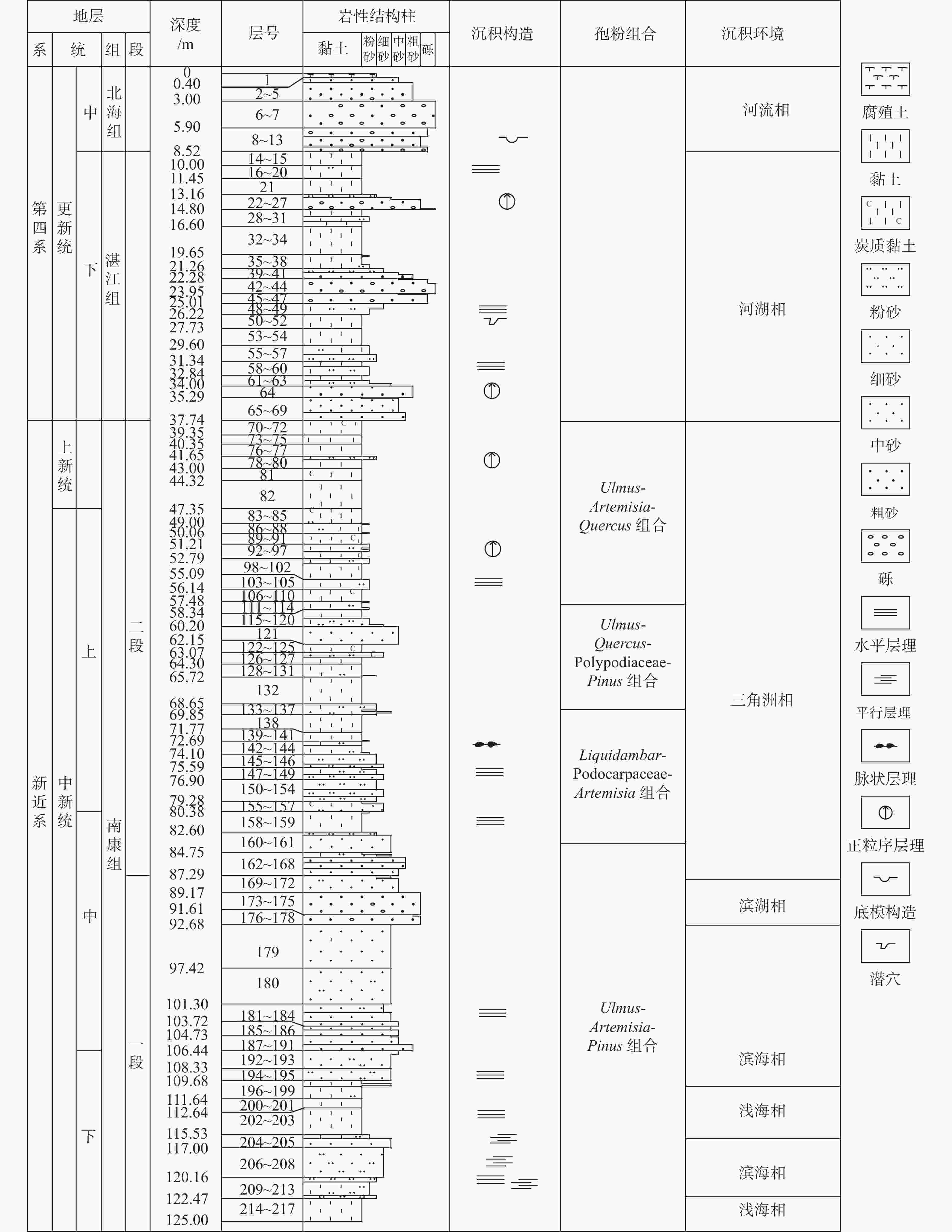
图 2 广西北海市ZKA02钻孔地层综合柱状图
Figure 2. Comprehensive stratigraphic column of Borehole ZKA02 in Beihai, Guangxi
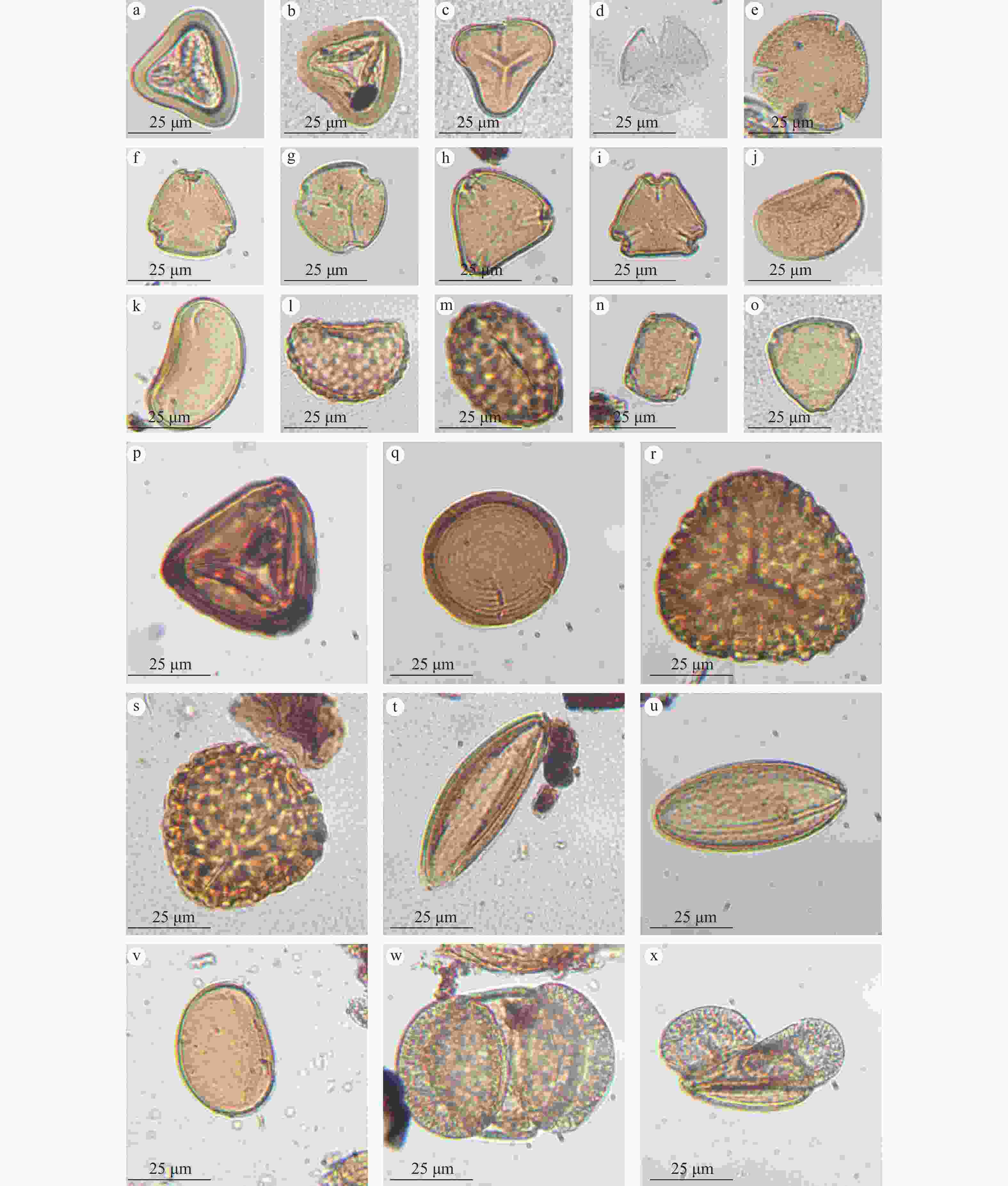
图 3 钻孔主要孢粉类型图片
a、b—凤尾蕨属;c—里白属;d、e—铃子香属;f—栾树属;g— 蔷薇属;h、i—山矾属;j、k—石韦属;l、m—水龙骨属;n—榆属;o—榛属;p—凤尾蕨属;q—环纹藻属;r、s—金粉蕨属;t、u—麻黄属;v—石韦属;w、x—松属
Figure 3. Photos of the main spore-pollen types in Borehole ZKA02
(a and b) Pteris; (c) Hicriopteris; (d and e) Chelonopsis; (f) Koelreuteria; (g) Rosa; (h and i) Symplocos; (j and k) Pyrrosia; (l and m) Polypodium; (n) Ulmus; (o) Corylus; (p) Pteris; (q) Concentricystes; (r and s) Onychiun; (t and u) Ephedra; (v) Pyrrosia; (w and x) Pinus
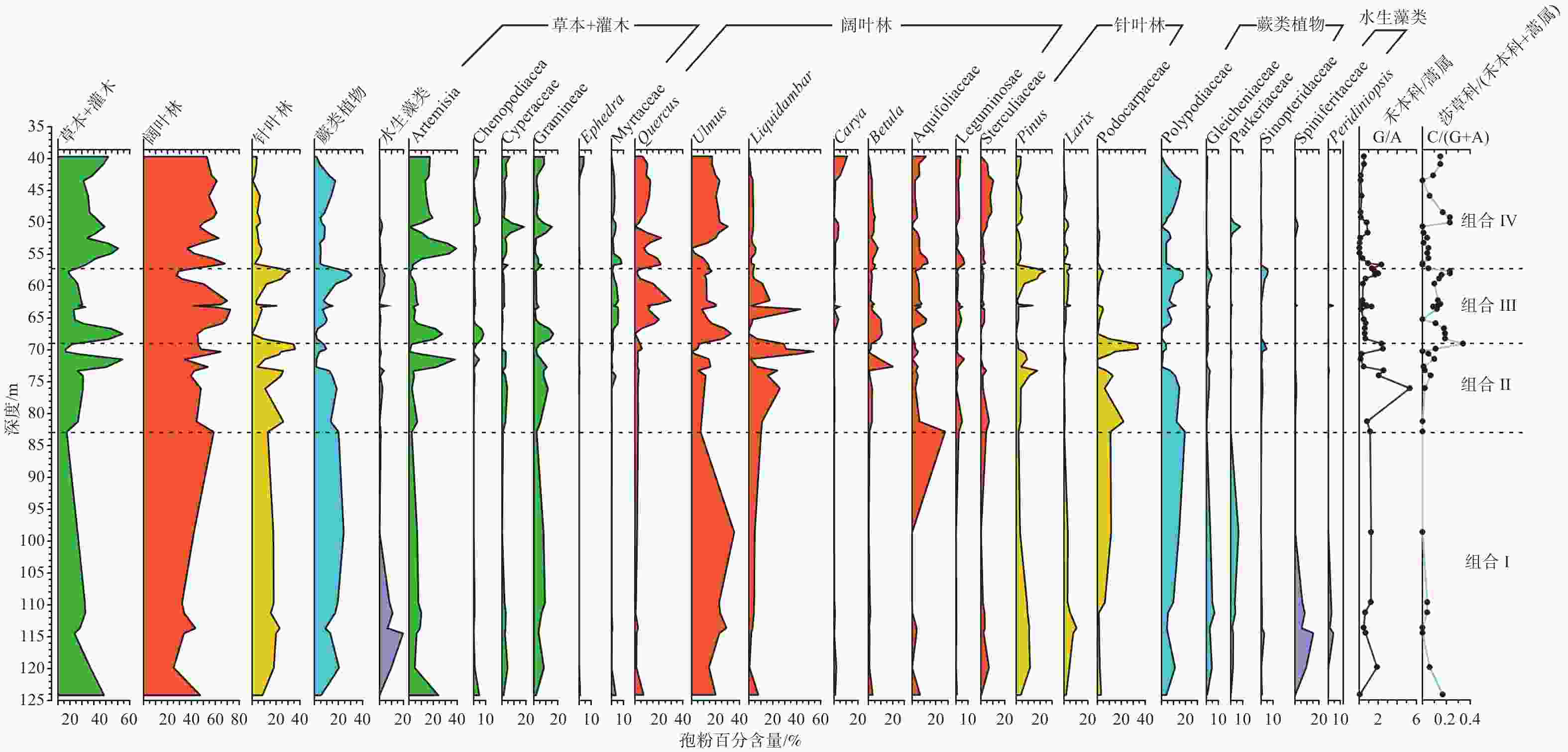
图 4 广西北海市ZKA02钻孔新生代地层孢粉百分比图谱与孢粉组合划分
Figure 4. Neogene spore-pollen percentage diagram and spore-pollen assemblages from Borehole ZKA02 in Beihai, Guangxi
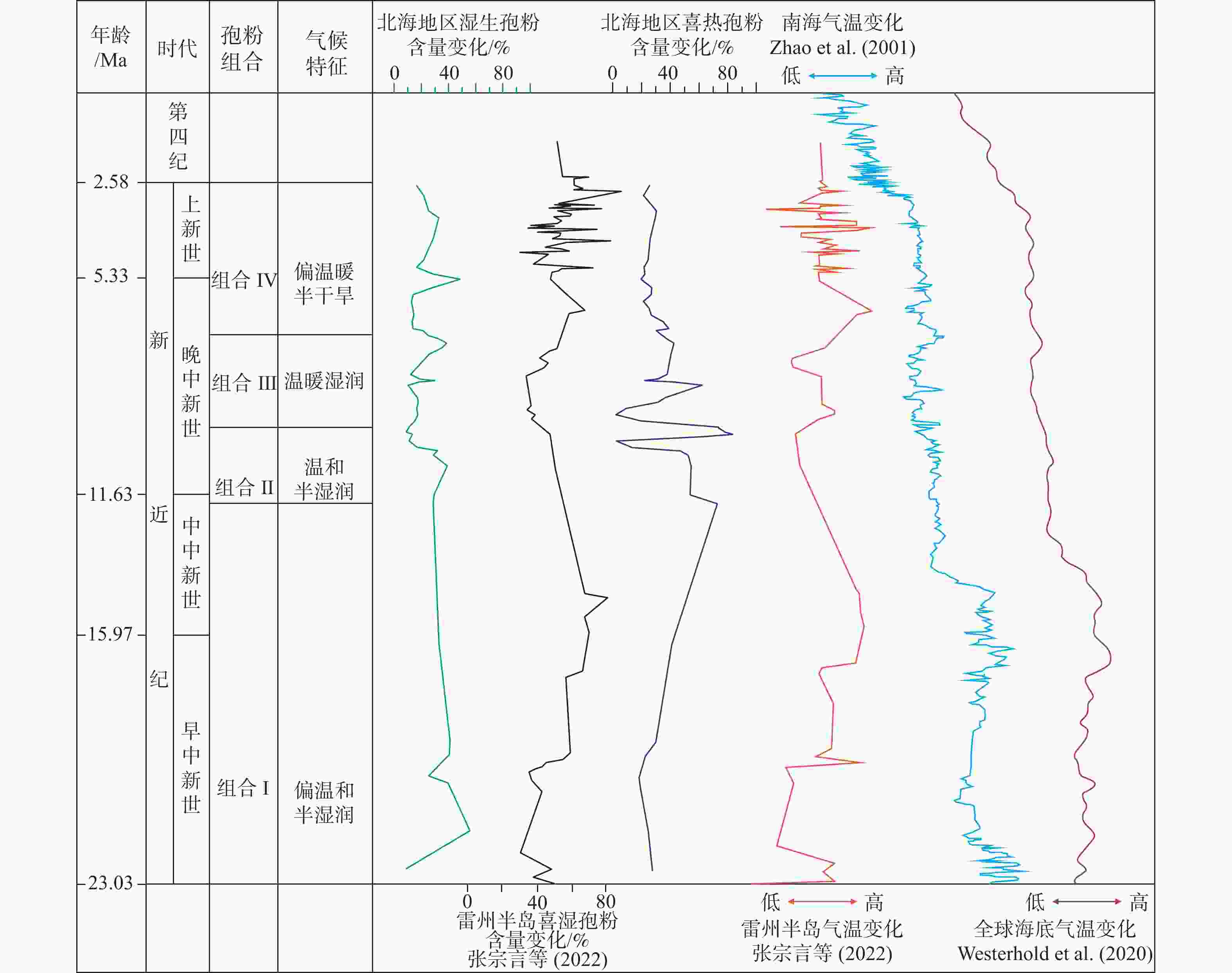
图 5 广西北海市ZKA02钻孔新近纪地层孢粉组合反映的气候变化与全球气候变化对比
Figure 5. The comparison between the climate change reflected by the Neogene spore-pollen assemblages in Borehole ZKA02, Beihai, Guangxi and the global climate change
表 1 北海地区及邻区新近纪岩石地层序列对比表
Table 1. Correlation of Neogene lithostratigraphic sequence of Beihai area and adjacent basins
 下载: 导出CSV
下载: 导出CSV
-
[1] Bureau of Geology and Mineral Resources of Guangxi Zhuang Autonomous Region, 1985. Regional geology of Guangxi Zhuang autonomous region[M]. Beijing: Geology Press. (in Chinese) [2] Bureau of Geology and Mineral Resources of Guangdong Province, 1996. Stratigraphy (lithostratic) of Guangdong province[M]. Wuhan: China University of Geosciences Press. (in Chinese) [3] Bureau of Geology and Mineral Resources of Guangxi Zhuang Autonomous Region, 1997. Stratigraphy (lithostratic) of Guangxi Zhuang autonomous region[M]. Wuhan: China University of Geosciences Press. (in Chinese) [4] CHEN P, WANG R, QIN J G, et al. , 2022. Oligocene-miocene palynostratigraphy and paleoclimate of well WN-A from deep water area, southeast Hainan Basin[J]. Journal of Jilin University (Earth Science Edition), 52(2): 390-402. (in Chinese with English Abstract) [5] CLIFT P D, 2020. Asian monsoon dynamics and sediment transport in SE Asia[J]. Journal of Asian Earth Sciences, 195: 104352. doi: 10.1016/j.jseaes.2020.104352 [6] DAI L, WENG C Y, LU J, et al. , 2012. Pollen and spore distribution in the surface sediments of the Northern South China Sea and their transportation[J]. Marine Geology & Quaternary Geology, 32(2): 93-108. (in Chinese with English Abstract) [7] DU J L, LI W Z, XU X M, et al. , 2009. Geological characteristics of Qianan formation, Kangbao county, Heibei province[J]. Geology and Mineral Resources of South China, 25(1): 14-19. (in Chinese with English Abstract) [8] EL-MOSLIMANY A P, 1990. Ecological significance of common nonarboreal pollen: examples from drylands of the Middle East[J]. Review of Palaeobotany and Palynology, 64(1-4): 343-350. doi: 10.1016/0034-6667(90)90150-H [9] FU N, LIN Q, WANG K, 2017. Main source rock reevaluation of Member 2 of Liushagang Formation in the sags of Beibuwan basin[J]. China Offshore Oil and Gas, 29(5): 12-21. (in Chinese with English Abstract) [10] Guangxi Institute of Geological Survey, 2017. Regional geology of Guangxi Zhuang autonomous region[R]. (in Chinese) [11] HERZSCHUH U, 2007. Reliability of pollen ratios for environmental reconstructions on the Tibetan Plateau[J]. Journal of Biogeography, 34(7): 1265-1273. doi: 10.1111/j.1365-2699.2006.01680.x [12] HUANG R H, HOU S G, MA C M, et al. , 2019. Review on palynological records in snow and ice[J]. Journal of Earth Sciences and Environment, 41(3): 362-378. (in Chinese with English Abstract) [13] HUANG X Q, CUI Z A, LIN H, et al. , 2022. The driving factors on climatic and palaeo-ecological evolution of Beibu gulf since Holocene[J]. Acta Geoscientica Sinica, 43(2): 129-143. (in Chinese with English Abstract) [14] JIANG Z X, ZENG L, LI M X, et al. , 1994. Tertiary in petroliferous regions of China (VIII): the north continental shelf region of South China Sea[M]. Beijing: Petroleum Industry Press. (in Chinese) [15] LEI H R, JIANG Z X, ZHOU H K, 2018. Analysis of paleoclimate evolution of the hyperthermal period in the early paleogene: taking the Dongying depression as an example[J]. Earth Science Frontiers, 25(4): 176-184. (in Chinese with English Abstract) [16] LI J, WANG R, QIN J G, et al. , 2020a. Paleogene-Neogene micropaleontological records of the Weixinan Depression, Beibuwan Basin and their paleoenvironmental significance[J]. Marine Geology & Quaternary Geology, 40(2): 29-36. (in Chinese with English Abstract) [17] LI J, ZHU Y H, QIN J G, 2020b. Late eocene to early miocene palynological assemblages of well WZ6-X in the southwest Weizhou sag, Beibuwan Basin, South China Sea and their paleoenvironment[J]. Acta Micropalaeontologica Sinica, 37(2): 175-187. (in Chinese with English Abstract) [18] LI X, ZHANG Z Y, ZHANG J Y, et al. , 2019. New progress in 1: 50 000 reginal survey in Leizhou peninsular area, Western Guangdong Province[J]. Geology and Mineral Resources of South China, 35(3): 293-305. (in Chinese with English Abstract) [19] LI Z, ZHANG Y L, LI Y X, et al. , 2010. Palynological records of Holocene monsoon change from the Gulf of Tonkin (Beibuwan), northwestern South China Sea[J]. Quaternary Research, 74(1): 8-14. doi: 10.1016/j.yqres.2010.04.012 [20] MAO X L, XU S L, LIU X Y, 2019. Late Cenozoic high resolution bio-stratigraphy and its bearing on sea-level fluctuation in the western Pearl River Mouth Basin[J]. Marine Geology & Quaternary Geology, 39(3): 40-50. (in Chinese with English Abstract) [21] MILLER K G, MOUNTAIN G S, 1996. Drilling and dating new jersey Oligocene-Miocene sequences: ice volume, global sea level, and exxon records[J]. Science, 271(5252): 1092-1095. doi: 10.1126/science.271.5252.1092 [22] Nanhai Branch of Petroleum Corporation of the People's Republic of China, Guangzhou Branch of Geological Exploration Company of the China, Editorial Board of the Tertiary of north continental shelf of South China Sea, 1981. Tertiary of north continental shelf of South China Sea[M]. Guangzhou: Guangdong Science and Technology Press. (in Chinese) [23] PAN A D, CHEN B S, LIU H P, et al. , 2008. Numerical techniques of spore-pollen data analysis on paleoclimate reconstruction[J]. Tropical Geography, 28(6): 493-497. (in Chinese with English Abstract) [24] PEI J L, ZHAO Y, ZHOU Z Z, et al. , 2021. Impact of Cenozoic Antarctic continent-ocean configuration patterns on global climate change[J]. Journal of Geomechanics, 27(5): 867-879. (in Chinese with English Abstract) [25] SHEN H, ZHAO K L, GE J Y, et al. , 2021. Early and Middle Pleistocene vegetation and its impact on faunal succession on the Liaodong Peninsula, Northeast China[J]. Quaternary International, 591: 15-23. doi: 10.1016/j.quaint.2020.11.034 [26] South Sea Branch of Petroleum Corporation of the People’s Republic of China, China National Geological Exploration Corporation Guangzhou Branch, Nanjing Institute of Geology and Palaeontology, et al. , 1981. Tertiary palaeontology of North continental shelf of South China Sea[M]. Guangzhou: Guangdong Science and Technology Press. (in Chinese) [27] TADA R, ZHENG H B, CLIFT P D, 2016. Evolution and variability of the Asian monsoon and its potential linkage with uplift of the Himalaya and Tibetan Plateau[J]. Progress in Earth and Planetary Science, 3(1): 4. doi: 10.1186/s40645-016-0080-y [28] TANG L Y, SHEN C M, LI C H, et al. , 2009. Pollen-inferred vegetation and environmental changes in the central Tibetan Plateau since 8200 yr BP[J]. Science in China Series D: Earth Sciences, 52(8): 1104-1114. (in Chinese) doi: 10.1007/s11430-009-0080-5 [29] WAN S M, LI A C, STUUT J B W, et al. , 2007. Grain-size records at ODP site 1146 from the northern South China sea: implications on the east Asian monsoon evolution since 20 Ma[J]. Science in China Series D: Earth Sciences, 50(10): 1536-1547. doi: 10.1007/s11430-007-0082-0 [30] WANG M M, ZHOU B G, JI F J, et al. , 2009. Fault activity on the Hepu Basin section of the Hepu-Beiliu fault[J]. Seismology and Geology, 31(3): 470-487. (in Chinese with English Abstract) [31] WANG M Y, WANG H Y, ZHU Z X, et al. , 2021. Late Miocene-Pliocene Asian summer monsoon variability linked to both tropical Pacific temperature and Walker Circulation[J]. Earth and Planetary Science Letters, 561: 116823. doi: 10.1016/j.jpgl.2021.116823 [32] WANG W M, 1992. Palynofloristic changes in the neogene of South China[J]. Acta Micropalaeontologica Sinica, 9(1): 81-95. (in Chinese with English Abstract) [33] WANG W M, 2011. Remarks on some possible bias in reconstructing neogene vegetation and environment in China[J]. Quaternary Sciences, 31(4): 589-596. (in Chinese with English Abstract) [34] WELTJE G J, 1997. End-member modeling of compositional data: numerical-statistical algorithms for solving the explicit mixing problem[J]. Mathematical Geology, 29(4): 503-549. doi: 10.1007/BF02775085 [35] WENG C Y, SUN X J, CHEN Y S, 1993. Numerical characteristics of pollen assemblages of surface samples from the West Kunlun Mountains[J]. Acta Botanica Sinica, 35(1): 69-79. (in Chinese with English Abstract) [36] WESTERHOLD T, MARWAN N, DRURY A J, et al. , 2020. An astronomically dated record of Earth's climate and its predictability over the last 66 million years[J]. Science, 369(6509): 1383-1387. doi: 10.1126/science.aba6853 [37] WU C L, 1984. Nan Hai (The South China Sea) movement and development of Basins in the South China Sea[J]. Marine Science Bulletin, 3(2): 47-53. (in Chinese with English Abstract) [38] WU K Y, LIU Y L, HU D S, et al. , 2021. Types and evolution of faults in the east area of the Wushi Sag, Beibuwan Basin[J]. Journal of Geomechanics, 27(1): 52-62. (in Chinese with English Abstract) [39] WU Z J, 1976. Characteristics of sporopollen assemblage and its stratigraphic significance in Well Weiqian-1, Weizhou Island, Beibuwan Basin[R]. (in Chinese) [40] XIANG K, LIU G Y, WEI F H, et al. , 2020. Quaternary palynological record from the borehole CZ04 in the Anxiang Sag of the Dongting Basin and its palaeoenvironmental significance[J]. Geology and Mineral Resources of South China, 36(2): 104-116. (in Chinese with English Abstract) [41] XIE J Y, LI J, MAI W, et al. , 2012. Palynofloras and age of the Liushagang and Weizhou Formations in the Beibuwan Basin, South China Sea[J]. Acta Palaeontologica Sinica, 51(3): 385-394. (in Chinese with English Abstract) [42] XIE J Y, QIN J G, LI J, et al. , 2014. Palynological assemblages of the Oligocene and Pliocene in the Yinggehai-Qiongdongnan Basin of the Northern South China Sea and their paleoenvironmental implications[J]. Acta Micropalaeontologica Sinica, 31(1): 85-97. (in Chinese with English Abstract) [43] XIE X N, ZHANG C, REN J Y, et al. , 2011. Effects of distinct tectonic evolutions on hydrocarbon accumulation in northern and southern continental marginal basins of South China Sea[J]. Chinese Journal of Geophysics, 54(12): 3280-3291. (in Chinese with English Abstract) [44] XU J Y, ZHANG L Y, 1999. Genesis of Cenozoic basins in the eastern margin of Eurasia Plate: dextral pulling-apart[J]. Oil & Gas Geology, 20(3): 187-191. (in Chinese with English Abstract) [45] YANG R Y, 1996. Association and time of Eocene sporo-pollen in Hepu Basin, Guangxi[J]. Guangxi Geology, 9(3): 31-41. (in Chinese with English Abstract) [46] YANG S R, 1993. Paleocene palynology of the Shangyang Formation, Hepu Basin, Guangxi, China[J]. Acta Micropalaeontologica Sinica, 10(2): 213-222. (in Chinese with English Abstract) [47] YANG X Q, ZHENG S J, YAN Y G, et al. , 2022. Response of terrigenous flux to the uplift of the Tibetan Plateau and summer monsoon evolution during the Late Miocene-Pliocene in the northern South China Sea[J]. Quaternary Sciences, 42(3): 650-661. (in Chinese with English Abstract) [48] YANG Z J, XU J M, 2002. Advances in studies on relationship among pollen, vegetation and climate[J]. Acta Phytoecologica Sinica, 26(S1): 73-81. (in Chinese with English Abstract) [49] YANG Z N, 1982. Zonation of Tertiary predominant palynological assemblages (TPPA) of Eastern China[J]. Marine Geological Research, 2(4): 63-71. (in Chinese with English Abstract) [50] YE D Q, ZHONG X C, YAO Y M, et al. , 1993. Tertiary in petroliferous regions of China (I): introduction[M]. Beijing: Petroleum Industry Press. (in Chinese) [51] ZACHOS J C, DICKENS G R, ZEEBE R E, 2008. An early Cenozoic perspective on greenhouse warming and carbon-cycle dynamics[J]. Nature, 451(7176): 279-283. doi: 10.1038/nature06588 [52] ZHANG J Y, 1987. Geological report of intensive survey in Fucheng exploration area, Hepu County, Guangxi[R]. (in Chinese) [53] ZHANG J Y, TAO M H, WANG J Y, et al. , 2015. Stratigraphyic division and correlation in Well Xi-2X in Hepu Basin, Guangxi[C]//Proceedings of the abstracts of the second meeting of the ninth session of the Chinese paleontological society palynology branch. Guiyang: Palaeontological Society of China. (in Chinese) [54] ZHANG Q R, 2009. A preliminary summary of cretaceous neogene sporopollen fossil assemblage sequence in central South China[M]. Wuhan: China University of Geosciences Press. (in Chinese) [55] ZHANG Y F, LIU D S, ZHANG X H, 2017. Neogene palynological assemblages from Qiongdongnan Basin and their paleoclimatic implications[J]. Marine Geology & Quaternary Geology, 37(1): 93-101. (in Chinese with English Abstract) [56] ZHANG Z Y, LIU X, LI X, et al. , 2020. The Late Oligocene-Early Pleistocene sporopollen assemblages and paleovegetation succession of core ZkA01 in Hetou town, Leizhou Peninsula of Guangdong Province[J]. Geological Bulletin of China, 39(6): 880-892. (in Chinese with English Abstract) [57] ZHANG Z Y, LIU X, LI X, et al. , 2022. Palynological study of the Late Oligocene-Early Pleistocene in Leizhou Peninsula, Guangdong, China: coexistence analysis and paleoclimate reconstruction[J]. Earth Science Frontiers, 29(2): 303-316. (in Chinese with English Abstract) [58] ZHAO F, ZHU Y H, JI X K, et al. , 2017. Late Oligocene to Early Miocene palynological assemblages of well LW5 in the deep water area, pearl river mouth Basin, northern South China Sea[J]. Acta Micropalaeontologica Sinica, 34(2): 201-210. (in Chinese with English Abstract) [59] ZHAO H T, WANG L R, YUAN J Y, 2007. Origin and time of Qiongzhou strait[J]. Marine Geology & Quaternary Geology, 27(2): 33-40. (in Chinese with English Abstract) [60] ZHAO Q H, JIAN Z M, WANG J L, et al. , 2001. Neogene oxygen isotopic stratigraphy, ODP Site 1148, northern South China Sea[J]. Science in China Series D: Earth Sciences, 44(10): 934-942. doi: 10.1007/BF02907086 [61] ZHAO Y, LIU H Y, LI F R, et al. , 2012. Application and limitations of the Artemisia/Chenopodiaceae pollen ratio in arid and semi-arid China[J]. The Holocene, 22(12): 1385-1392. doi: 10.1177/0959683612449762 [62] ZHU R, ZHANG C M, DU J Y, et al. , 2015. Controls of neogene sea level change on sand bodies in the pearl river mouth basin[J]. Geological Journal of China Universities, 21(4): 685-693. (in Chinese with English Abstract) [63] ZHU Z Y, QIU Y, ZHOU H Y, et al. , 2002. A summary review of research progress on global change in the South China Sea[J]. Journal of Geomechanics, 8(4): 315-322, 314. (in Chinese with English Abstract) [64] 陈平, 王任, 覃军干, 等, 2022. 琼东南盆地深水区WN-A井渐新世—中新世孢粉地层学及古气候[J]. 吉林大学学报(地球科学版), 52(2): 390-402. [65] 戴璐, 翁成郁, 陆钧, 等, 2012. 南海北部表层沉积物的孢粉分布及其传播机制[J]. 海洋地质与第四纪地质, 32(2): 93-108. [66] 杜金利, 李卫忠, 徐旭明, 等, 2009. 河北康保县迁安组的沉积环境、孢粉组合及时代[J]. 华南地质与矿产, 25(1): 14-19. [67] 傅宁, 林青, 王柯, 2017. 北部湾盆地主要凹陷流沙港组二段主力烃源岩再评价[J]. 中国海上油气, 29(5): 12-21. [68] 广东省地质矿产局, 1996. 广东省岩石地层[M]. 武汉: 中国地质大学出版社. [69] 广西壮族自治区地质调查研究院, 2017. 广西壮族自治区区域地质志[R]. [70] 广西壮族自治区地质矿产局, 1985. 广西壮族自治区区域地质志[M]. 北京: 地质出版社. [71] 广西壮族自治区地质矿产局, 1997. 广西壮族自治区岩石地层[M]. 武汉: 中国地质大学出版社. [72] 黄壬晖, 侯书贵, 马春梅, 等, 2019. 雪冰孢粉记录研究进展[J]. 地球科学与环境学报, 41(3): 362-378. [73] 黄向青, 崔振昂, 林海, 等, 2022. 北部湾全新世气候与古生态环境演进特征及其驱动因素[J]. 地球学报, 43(2): 129-143. [74] 蒋仲雄, 曾麟, 李明兴, 等, 1994. 中国油气区第三系(VIII): 南海北部大陆架油气区分册[M]. 北京: 石油工业出版社. [75] 雷华蕊, 姜在兴, 周红科, 2018. 早古近纪极热时期古气候演化分析: 以东营凹陷为例[J]. 地学前缘, 25(4): 176-184. [76] 李君, 王任, 覃军干, 等, 2020a. 北部湾盆地涠西南凹陷古、新近系古生物记录及古环境意义[J]. 海洋地质与第四纪地质, 40(2): 29-36. [77] 李君, 祝幼华, 覃军干, 2020b. 南海北部湾盆地涠西南凹陷WZ6-X井晚始新世—早中新世孢粉组合及其古环境[J]. 微体古生物学报, 37(2): 175-187. [78] 李响, 张宗言, 张楗钰, 等, 2019. 粤西雷州半岛地区1∶5万区域地质调查成果与主要进展[J]. 华南地质与矿产, 35(3): 293-305. [79] 毛雪莲, 徐守立, 刘新宇, 2019. 珠江口盆地西部新近纪高分辨率生物地层及海平面变化分析[J]. 海洋地质与第四纪地质, 39(3): 40-50. [80] 潘安定, 陈碧姗, 刘会平, 等, 2008. 孢粉学定量重建古气候方法探讨[J]. 热带地理, 28(6): 493-497. [81] 裴军令, 赵越, 周在征, 等, 2021. 南极新生代海陆格局变迁对全球气候变化的影响[J]. 地质力学学报, 27(5): 867-879. [82] 唐领余, 沈才明, 李春海, 等, 2009. 花粉记录的青藏高原中部中全新世以来植被与环境[J]. 中国科学(D辑: 地球科学), 39(5): 615-625. [83] 万世明, 李安春, STUUT J B W, 等, 2007. 南海北部ODP1146站粒度揭示的近20Ma以来东亚季风演化[J]. 中国科学 D辑: 地球科学, 37(6): 761-770. [84] 王明明, 周本刚, 计凤桔, 等, 2009. 合浦-北流断裂带西支合浦盆地段断裂活动性研究[J]. 地震地质, 31(3): 470-487. [85] 王伟铭, 1992. 中国南方晚第三纪孢粉植物群的变迁[J]. 微体古生物学报, 9(1): 81-95. [86] 王伟铭, 2011. 中国新近纪植被和环境研究中的几个问题讨论[J]. 第四纪研究, 31(4): 589-596. [87] 翁成郁, 孙湘君, 陈因硕, 1993. 西昆仑地区表土花粉组合特征及其与植被的数量关系[J]. 植物学报, 35(1): 69-79. [88] 吴长林, 1984. 南海运动与南海诸盆地的发育[J]. 海洋通报, 3(2): 47-53. [89] 吴孔友, 刘煜磊, 胡德胜, 等, 2021. 北部湾盆地乌石凹陷东区断裂类型及其形成演化[J]. 地质力学学报, 27(1): 52-62. [90] 吴作基, 1976. 北部湾涠洲岛涠浅一井的孢粉组合特征及其地层意义[R]. [91] 向轲, 刘庚寅, 魏方辉, 等, 2020. 洞庭盆地安乡凹陷CZ04孔第四纪孢粉记录及其古环境意义[J]. 华南地质与矿产, 36(2): 104-116. [92] 谢金有, 李君, 麦文, 等, 2012. 北部湾盆地涠洲组和流沙港组孢粉组合及时代[J]. 古生物学报, 51(3): 385-394. [93] 谢金有, 覃军干, 李君, 等, 2014. 南海莺琼盆地渐新世—上新世孢粉组合序列及其古环境意义[J]. 微体古生物学报, 31(1): 85-97. [94] 解习农, 张成, 任建业, 等, 2011. 南海南北大陆边缘盆地构造演化差异性对油气成藏条件控制[J]. 地球物理学报, 54(12): 3280-3291. [95] 许浚远, 张凌云, 1999. 欧亚板块东缘新生代盆地成因: 右行剪切拉分作用[J]. 石油与天然气地质, 20(3): 187-191. [96] 杨荣玉, 1996. 广西合浦盆地下第三系孢粉组合及时代[J]. 广西地质, 9(3): 31-41. [97] 杨世蓉, 1993. 广西合浦盆地古新世上洋组孢粉组合及其地层意义[J]. 微体古生物学报, 10(2): 213-222. [98] 杨小强, 郑思静, 闫永刚, 等, 2022. 南海北部晚中新世-上新世期间碎屑物质输入对青藏高原隆升与季风演化的响应[J]. 第四纪研究, 42(3): 650-661. doi: 10.11928/j.issn.1001-7410.2022.03.03 [99] 杨振京, 徐建明, 2002. 孢粉-植被-气候关系研究进展[J]. 植物生态学报, 26(S1): 73-81. [100] 杨祖年, 1982. 广东第三系孢粉优势组合带与我国东部各盆地的对比[J]. 海洋地质研究, 2(4): 63-71. [101] 叶得泉, 钟筱春, 姚益民, 等, 1993. 中国油气区第三系(I): 总论[M]. 北京: 石油工业出版社. [102] 张家渝, 1987. 广西壮族自治区合浦县合浦煤田福成探区精查地质报告[R]. [103] 张金岩, 陶明华, 王婧雅, 等, 2015. 广西合浦盆地西2X井地层划分对比研究[C]//中国古生物学会孢粉学会第九届二次学术年会论文摘要集. 贵阳: 中国古生物学会. [104] 张清如, 2009. 中南地区白垩纪—新近纪孢粉化石组合序列的初步总结[M]. 武汉: 中国地质大学出版社. [105] 张一凡, 刘东生, 张训华, 2017. 琼东南盆地新生代孢粉组合及其古气候意义[J]. 海洋地质与第四纪地质, 37(1): 93-101. [106] 张宗言, 刘祥, 李响, 等, 2020. 广东雷州半岛河头镇ZKA01钻孔剖面晚渐新世—早更新世孢粉组合及古植被演替[J]. 地质通报, 39(6): 880-892. [107] 张宗言, 刘祥, 李响, 等, 2022. 广东雷州半岛晚渐新世—早更新世孢粉共存因子分析及古气候变化重建[J]. 地学前缘, 29(2): 303-316. doi: 10.13745/j.esf.sf.2022.2.9 [108] 赵飞, 祝幼华, 季兴开, 等, 2017. 南海北部珠江口盆地深水区LW5井晚渐新世—早中新世孢粉组合[J]. 微体古生物学报, 34(2): 201-210. [109] 赵焕庭, 王丽荣, 袁家义, 2007. 琼州海峡成因与时代[J]. 海洋地质与第四纪地质, 27(2): 33-40. [110] 中华人民共和国石油公司南海分公司, 中华人民共和国地质勘探公司广州分公司, 《南海北部大陆架第三系》编委会, 1981. 南海北部大陆架第三系[M]. 广州: 广东科技出版社. [111] 中华人民共和国石油勘探公司南海分公司, 中华人民共和国地质勘探公司广州分公司, 中国科学院南京地质古生物研究所, 等, 1981. 南海北部大陆架第三纪古生物图册[M]. 广州: 广东科技出版社. [112] 朱锐, 张昌民, 杜家元, 等, 2015. 珠江口盆地新近纪海平面升降过程及其对砂体的控制[J]. 高校地质学报, 21(4): 685-693. [113] 朱照宇, 邱燕, 周厚云, 等, 2002. 南海全球变化研究进展[J]. 地质力学学报, 8(4): 315-322, 314. -
 下载:
下载:





计量
- 文章访问数: 1261
- HTML全文浏览量: 315
- PDF下载量: 58
- 被引次数: 0
