

The spore-pollen assemblages of the Miocene Baode Formation from Borehole ZK301 in the Yuncheng Basin and its geological significance
-
摘要:
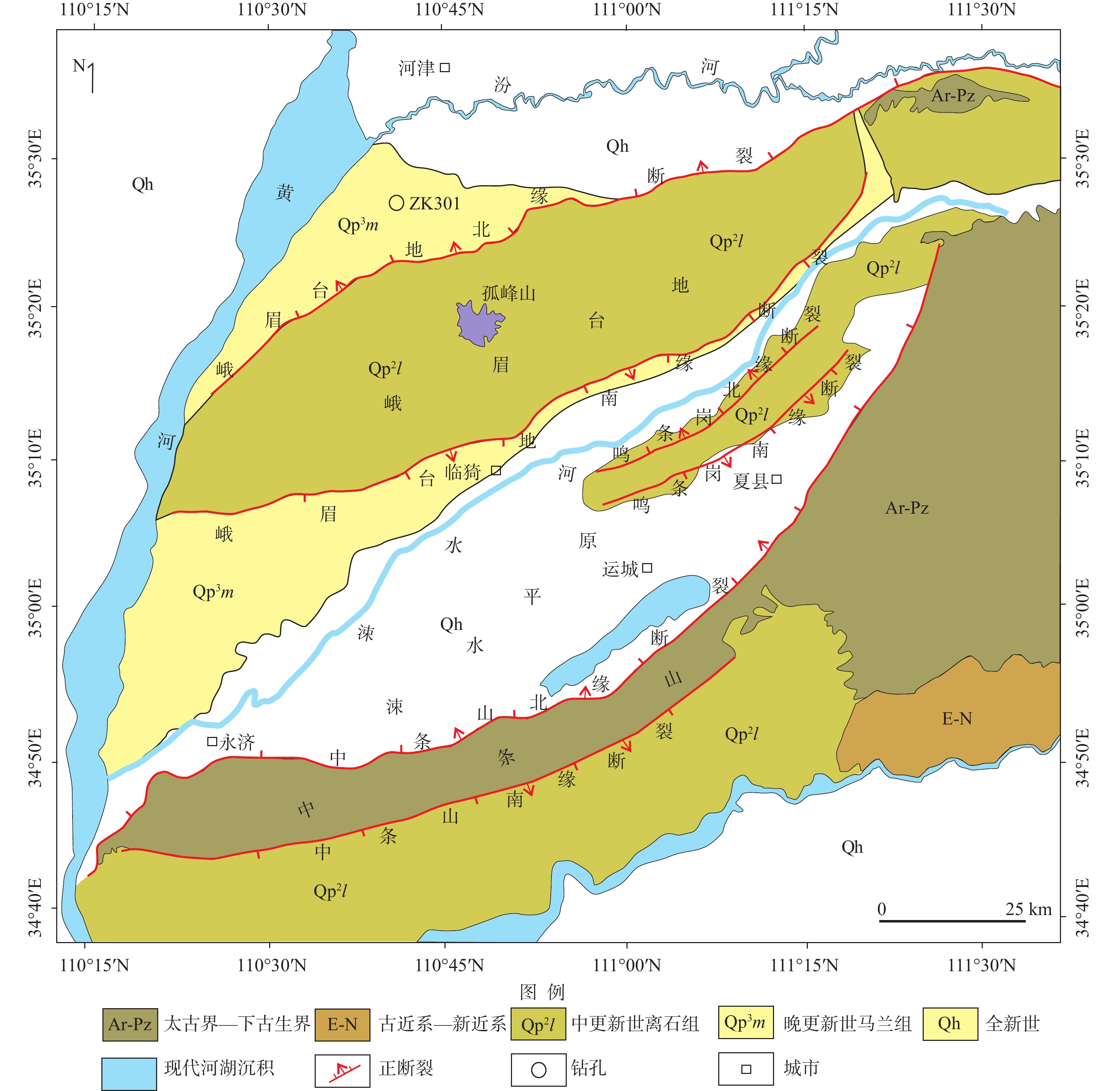
中新世保德组沉积时期是青藏高原向北东方向隆升扩展影响到鄂尔多斯周缘新生代盆地的关键时期。文章对运城盆地峨嵋台地ZK301钻孔中新世晚期保德组岩芯进行了孢粉学研究,系统鉴定了沉积孢粉和再沉积孢粉的特征,探讨了保德组沉积时期的古气候与古构造背景。运城盆地中新世保德组沉积孢粉自下而上可以划分为麻黄科−藜科−禾本科和蒿属−藜科−葎草属2个孢粉组合带,说明运城盆地中新世晚期是从以麻黄科−藜科−禾本科为主的荒漠草原发展为以蒿属−藜科为主的荒漠草原,气候由相对寒冷干燥过渡为偏冷偏干,这种变化过程与中新世晚期青藏高原隆升扩展远程效应对气候的影响有关。再沉积孢粉组合主要集中在保德组下部,以麻黄科含量最高,其次是松属、云杉属、柏科、藜科、胡桃科和蕨类植物孢子,还有少量的克拉梭粉属和胡颓子科等,反映了温暖湿润的气候环境。再沉积孢粉主要来自于中条山北缘山前的古近纪地层,说明在保德组沉积初期,中条山存在一期快速的隆升剥蚀,保德组经历了古近纪地层的剥蚀−搬运−再沉积过程。
Abstract:The deposition period of the Baode Formation in the Miocene was crucial when the uplift and expansion of the Tibetan Plateau in the NE direction affected the Cenozoic basins around the Ordos Plateau. Previous research has mainly focused on tectonic and sedimentary aspects, with relatively few results on climate and environmental responses. We conducted a systematic sporopollen study on the Borehole ZK301 from the late Miocene Baode Formation in the E’mei tableland, Yuncheng Basin, characterized the deposited and redeposited spore-pollen and discussed the paleoclimate and paleotectonic background of the Baode Formation during its deposition. From bottom to top, the Miocene Baode Formation in the Yuncheng Basin can be divided into two spore-pollen assemblages, which are Ephedraceae–Chenopodiaceae–Gramineae zone and Artemisia–Chenopodiaceae–Humulus zone, indicating that the desert steppe dominated by Chenopodiaceae, Gramineae, and Ephedraceae developed into the desert steppe dominated by Artemisia and Chenopodiaceae in the late Miocene. Accordingly, the climate transitioned from relatively cold and dry to cold and dry, which was related to the influence of the remote effect of the uplift and expansion of the Tibetan Plateau on the climate in the late Miocene. The redeposited spore-pollen assemblages are mainly concentrated in the lower part of the Baode Formation, with the highest content of Ephedraceae, followed by Pinus, Picea, Cupressaceae, Chenopodiaceae, Juglandaceae and Pteridophyte, and a small amount of Classopollis and Elaeagnaceae, reflecting a warm and humid climate. The redeposited spore-pollens were mainly from the Paleogene strata on the northern margin of the Zhongtiaoshan Mountains, which indicates that there was a rapid uplift and denudation of the Zhongtiaoshan Mountains in the early stage of the Baode Formation deposition. The Baode Formation underwent the Paleogene strata’s denudation, transportation, and redeposited process. The research results can provide evidence for the late Miocene paleoclimate in the Yuncheng Basin and new evidence for the uplift of the Zhongtiaoshan Mountains in this period.
-
Key words:
- Baode Formation /
- palynological records /
- paleoclimate /
- tectonic implications /
- Yuncheng Basin /
- Cenozoic
-
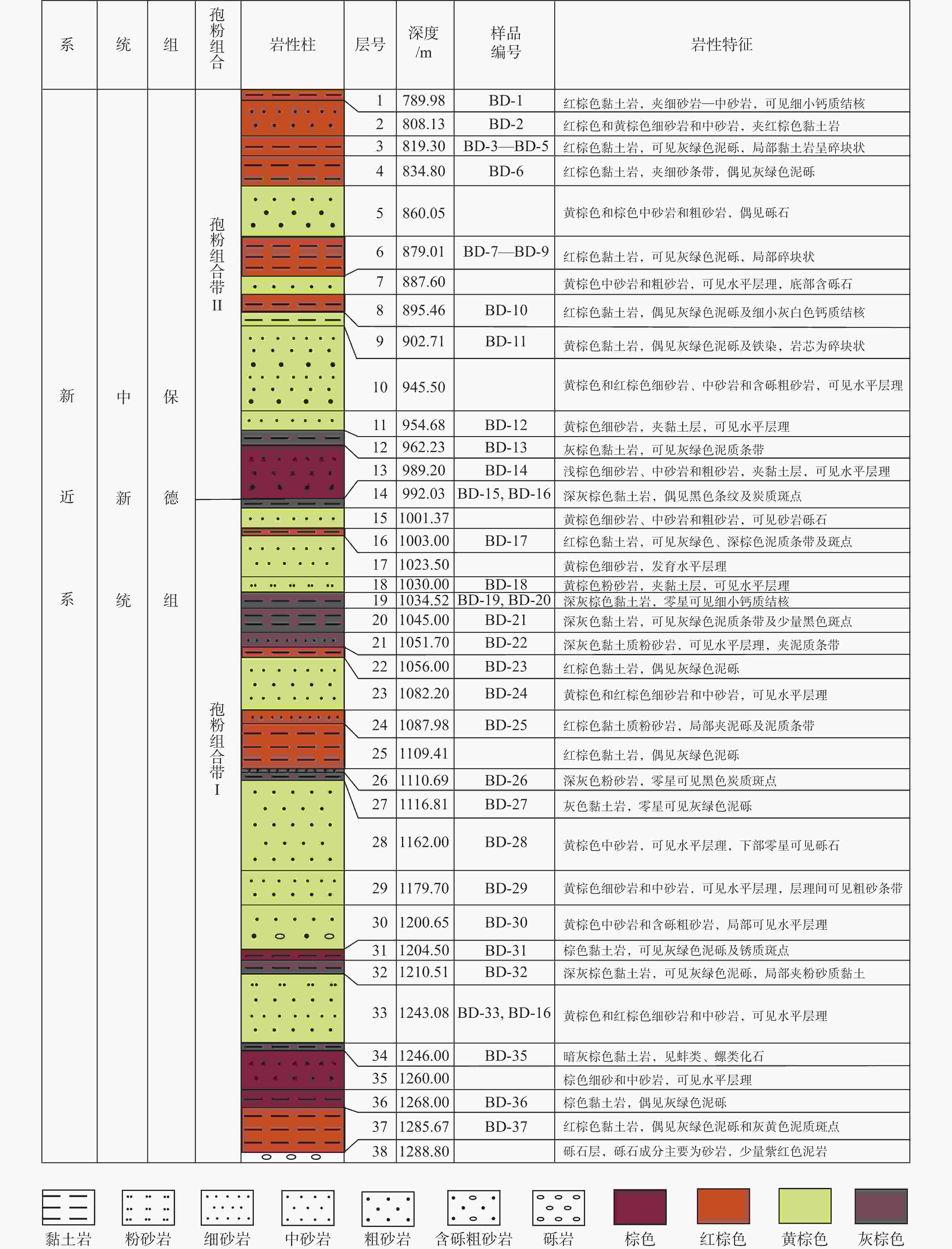
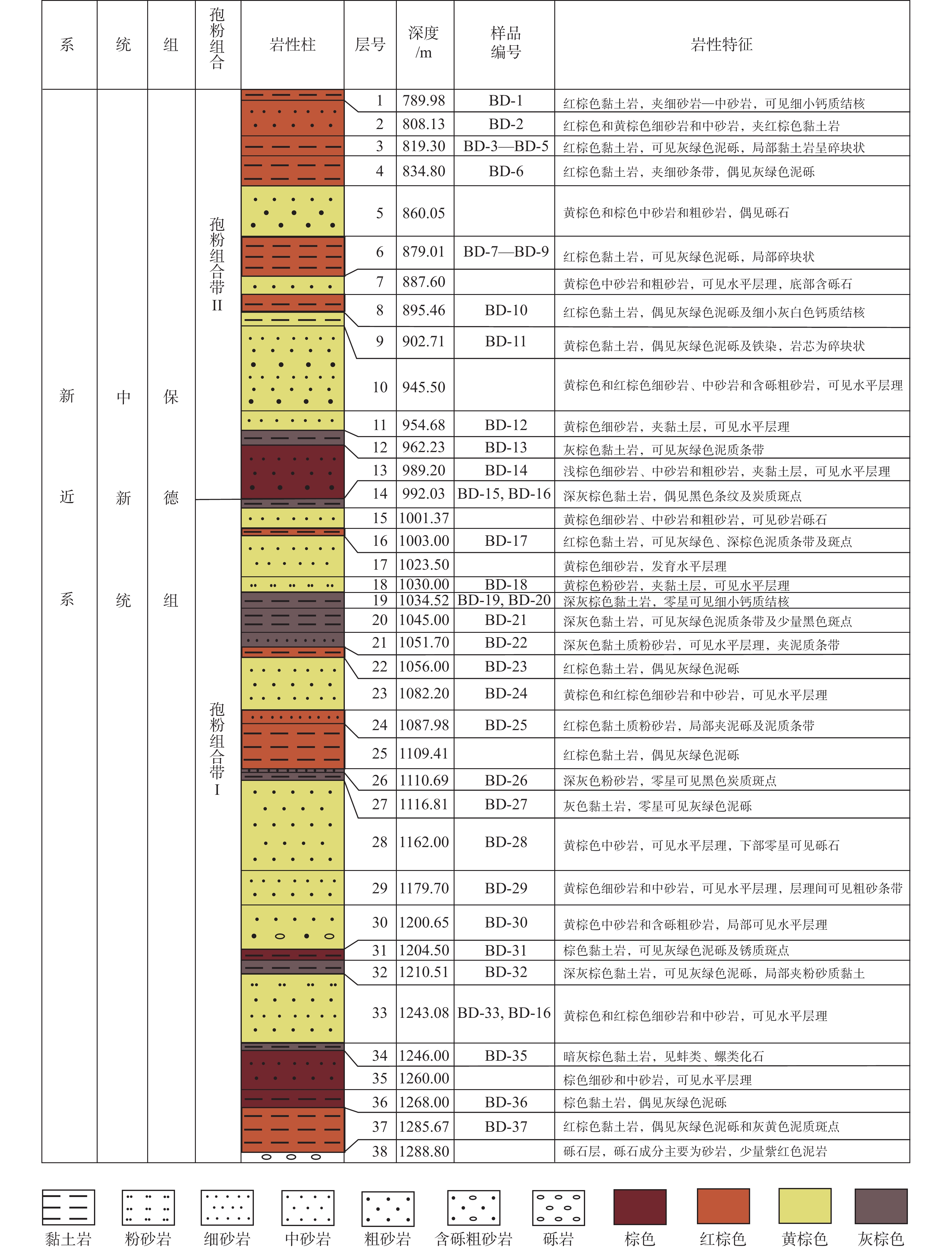
图 2 ZK301钻孔中新世晚期保德组剖面柱状图
Figure 2. Histogram of the late Miocene Baode Formation from the Borehole ZK301
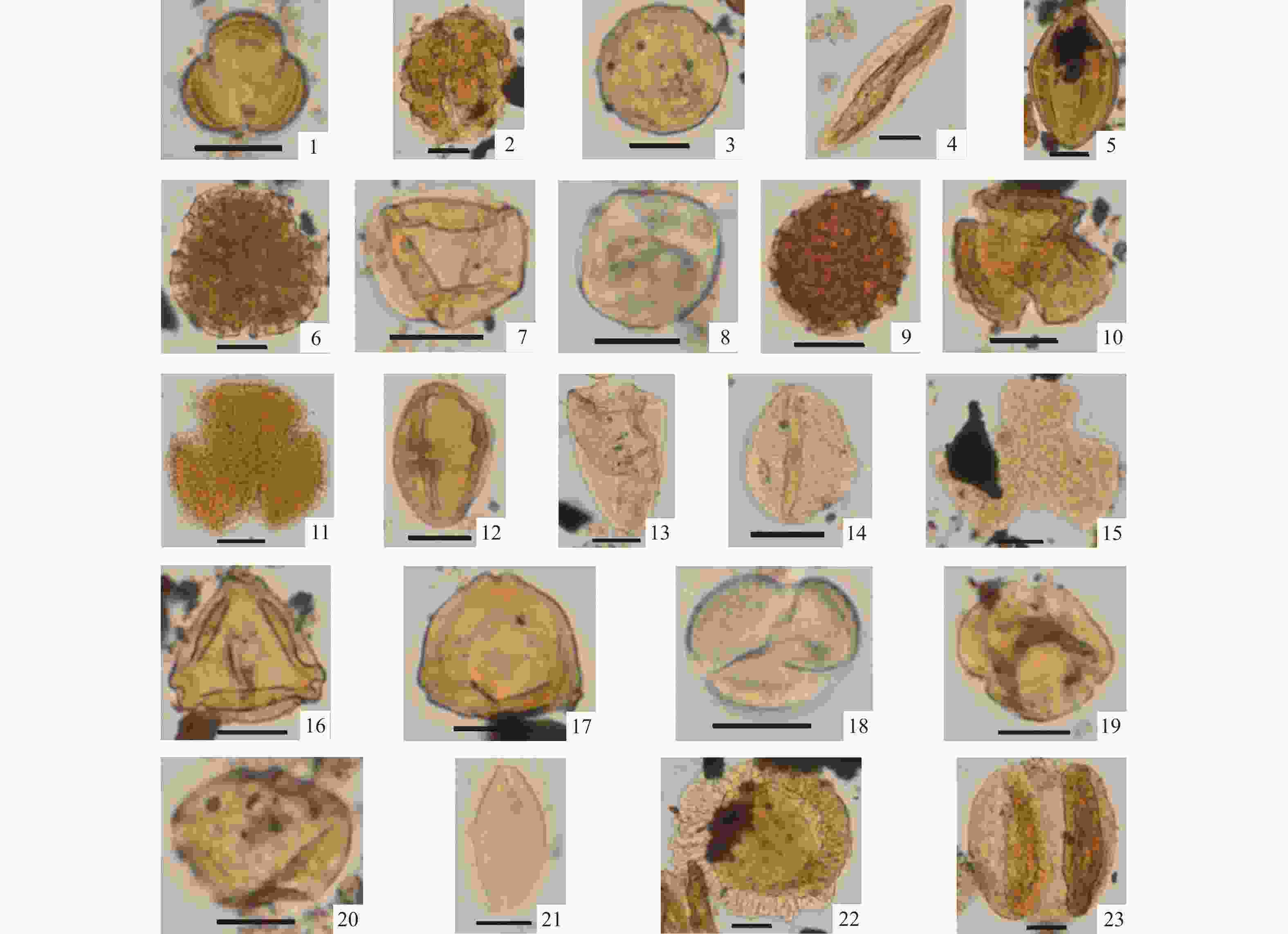
图 3 ZK301钻孔中新世晚期保德组主要孢粉类型(比例尺均为10 μm)
1—蒿属;2—蓝刺头属;3—藜科;4—麻黄科;5—白刺属;6—白花丹科;7—禾本科;8—葎草属;9—蓼科;10—毛茛科;11—牻牛儿苗科;12—伞形科;13—莎草科;14—眼子菜科;15—槭树科;16—桦木科;17—胡桃科;18—柳属;19—椴树科;20—榆科;21—苏铁科;22—铁杉属;23—松属
Figure 3. Photomicrographs of selected spore-pollen types from the late Miocene Baode Formation from Borehole ZK301 (The scale is 10 μm)
(1) Artemisia; (2) Echinops; (3) Chenopodiaceae; (4) Ephedraceae; (5) Nitraria; (6) Plumbaginaceae; (7) Gramineae; (8) Humulus; (9) Polygonaceae; (10) Ranunculaceae; (11) Geraniaceae; (12) Apiaceae; (13) Cyperaceae; (14) Potamogetonaceae; (15) Aceraceae; (16) Betulaceae; (17) Juglandaceae; (18) Salix; (19) Tiliaceae; (20) Ulmaceae; (21) Cycadaceae; (22) Tsuga; (23) Pinus
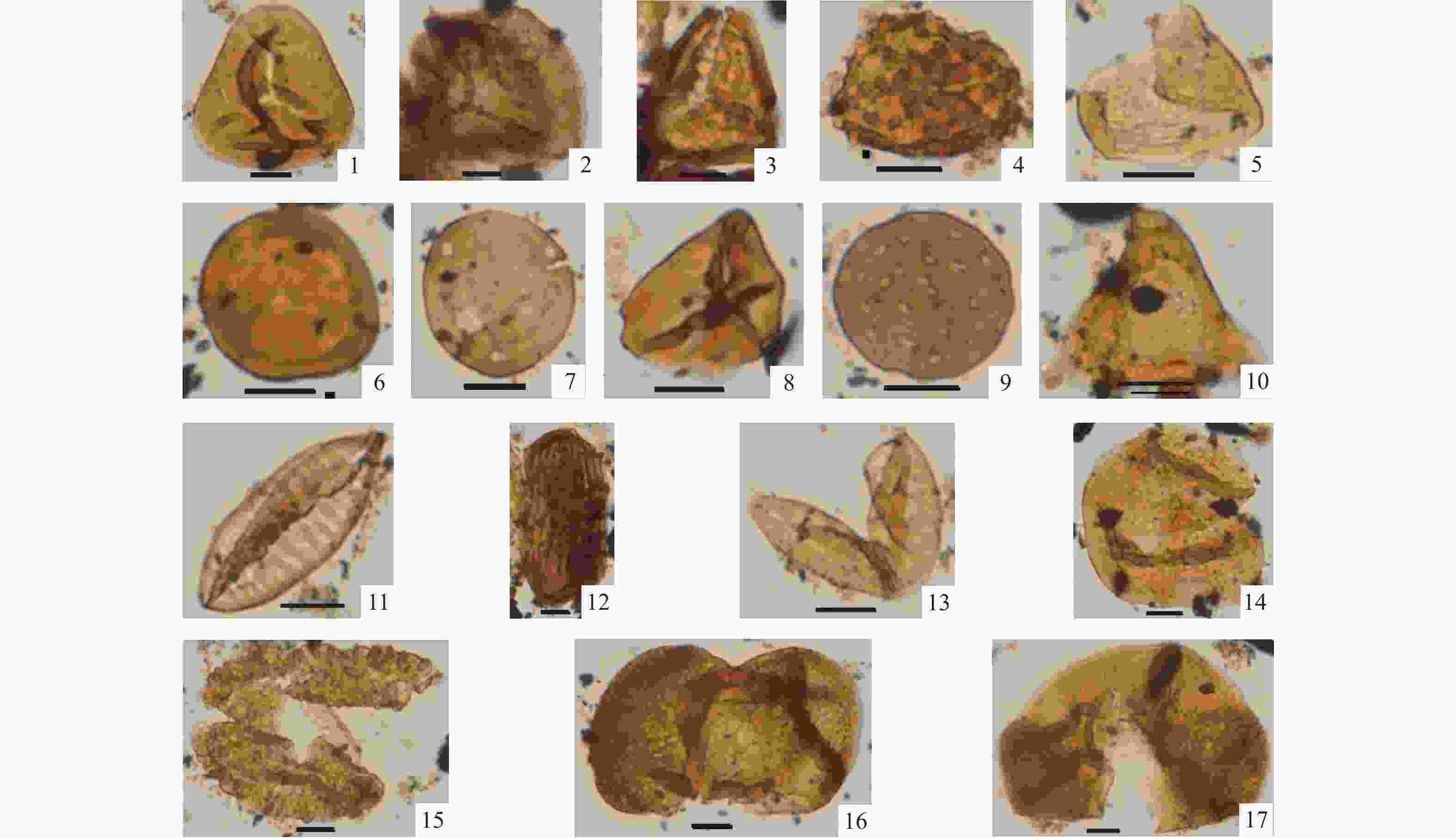
图 4 ZK301钻孔中新世晚期保德组主要再沉积孢粉类型(比例尺均为10 μm)
1—里白科;2—凤尾蕨科;3—卷柏科;4—紫萁科;5—水蕨科;6—克拉梭粉属;7—胡桃科:山核桃属;8—胡桃科:黄杞属;9—藜科;10—胡颓子科;11—麻黄科:梭形麻黄粉;12—麻黄科:多肋麻黄粉;13—柏科;14—南洋杉科;15—铁杉属;16—松属;17—云杉属
Figure 4. Photomicrographs of selected redeposited spore-pollen types of the late Miocene Baode Formation from Borehole ZK301 (The scale is 10 μm)
(1) Gleicheniaceae; (2) Pteridaceae; (3) Selaginellaceae; (4) Osmundaceae; (5) Parkeriaceae; (6) Classopollis; (7) Juglandaceae: Carya; (8) Juglandaceae: Engelhardtia; (9) Chenopodiaceae; (10) Elaeagnaceae; (11) Ephedraceae: Ephedripites fusiformis; (12) Ephedraceae: Ephedripites multicotatus; (13) Cupressaceae; (14) Araucariaceae; (15) Tsuga; (16) Pinus; (17) Picea
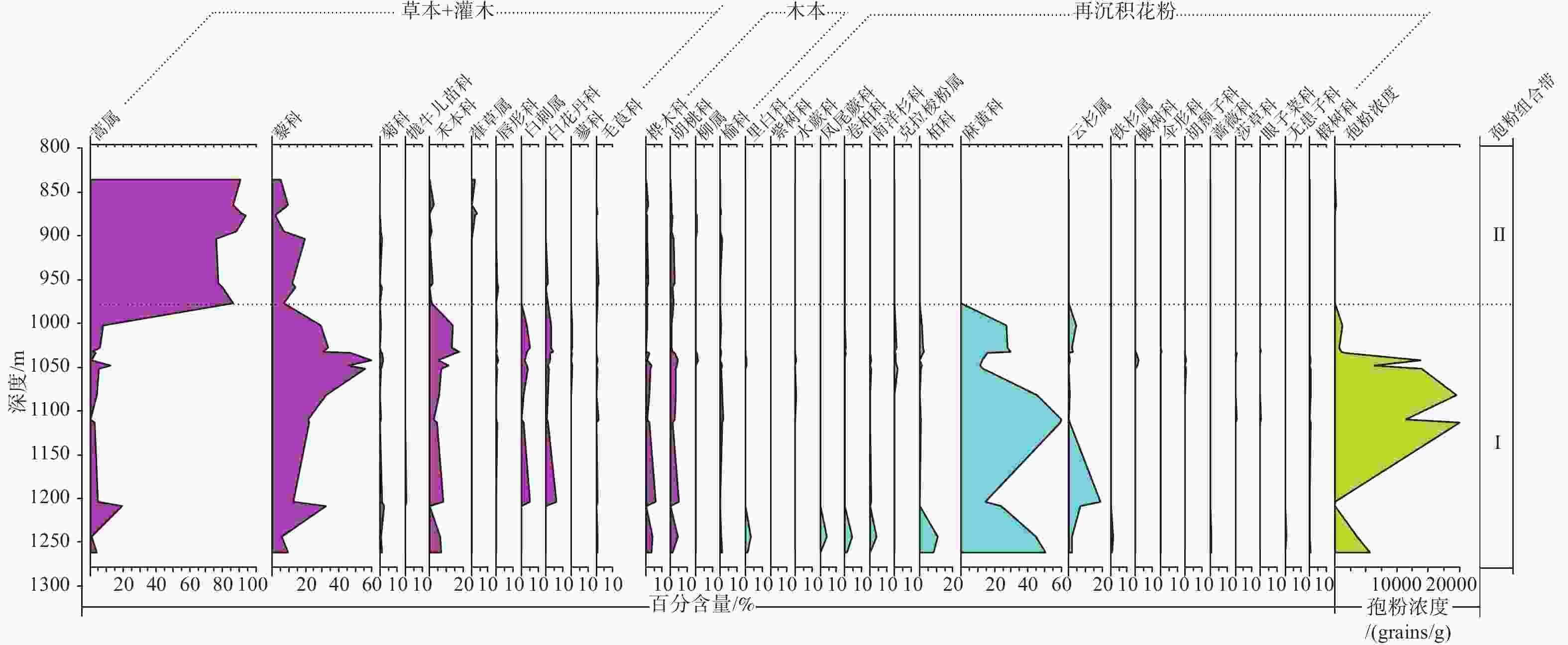
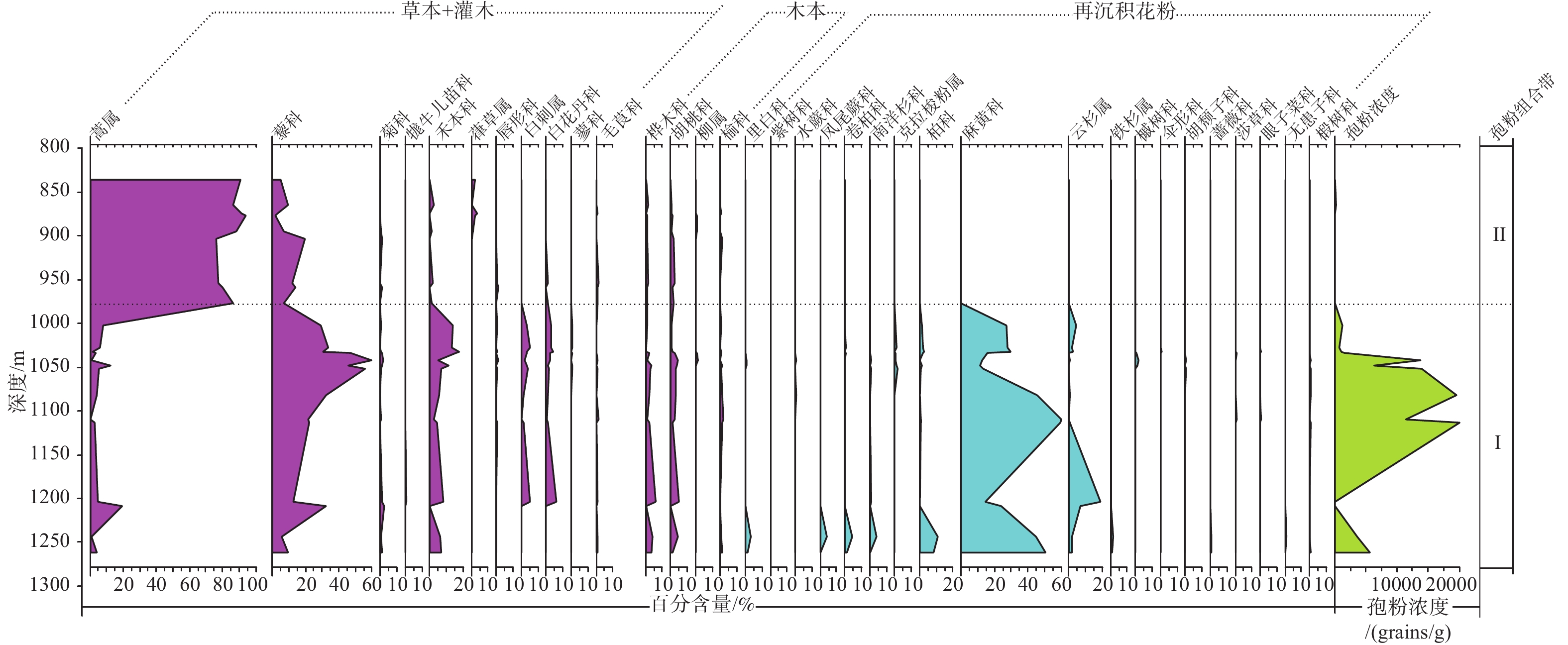
图 5 ZK301钻孔中新世晚期保德组孢粉百分比图谱
Figure 5. Spore-pollen percentage diagram of the late Miocene Baode Formation from Borehole ZK301
-
[1] CHEN X Q, 2017. Sedimentary and tectonic evolution of the Cenozoic Weihe-Shanxi Graben and the mechanism [D]. Doctor's thesis. Beijing: Peking University. (in Chinese with English abstract) [2] CHEN X Q, DONG S W, SHI W, et al. , 2021. Magnetostratigraphic ages of the Cenozoic Weihe and Shanxi Grabens in North China and their tectonic implications[J]. Tectonophysics, 813: 228914. doi: 10.1016/j.tecto.2021.228914 [3] China Vegetation Editing Committee, 1980. China vegetation[M]. Beijing: Science Press. (in Chinese) [4] DENG T, WANG W M, YUE L P, et al. , 2004. New advances in the establishment of the Neogene Baode stage[J]. Journal of Stratigraphy, 28(1): 41-47. (in Chinese with English abstract) [5] DENG T, WANG W M, YUE L P, 2008. A comprehensive study report on the Shanwang and Baode stages of the Neogene in China[M]. In: Third National Stratigraphic Commission, ed. Research Report on the Stratigraphic Formation of Major Faults in China (2001~2005). Beijing: Geology Press, 13-31. (in Chinese) [6] FANG X M, 2017. Phased uplift of the Tibetan Plateau[J]. Science & Technology Review, 35(6): 42-50. (in Chinese with English abstract) [7] GRUBOV V I, 2001. Key to the vascular plants of Mongolia (with an Atlas)[M]. Science Publishers. [8] HAN X M, LIU F, ZHANG W T, et al. , 2015. Analyzing the variation characteristics of stress field in Hetao seismic belt using focal mechanism data[J]. Seismology and Geology, 37(4): 1030-1042. (in Chinese with English abstract) [9] KOU L L, LI Z H, DONG X P, et al. , 2021. The age sequence of the detrital zircons from the Guanyindian section in Longde, the northeastern margin of the Tibetan Plateau, and its geological significance[J]. Journal of Geomechanics, 27(6): 1051-1064. (in Chinese with English abstract) [10] LI S Z, CAO X Z, WANG G Z, et al. , 2019. Meso-Cenozoic tectonic evolution and plate reconstruction of the Pacific Plate[J]. Journal of Geomechanics, 25(5): 642-677. (in Chinese with English abstract) [11] LI W Y, 1998. Quaternary vegetation and environment of China[M]. Beijing: Science Press. (in Chinese) [12] LI X R, FANG X M, WU F L, et al. , 2011. Pollen evidence from Baode of the northern Loess Plateau of China and strong East Asian summer monsoons during the Early Pliocene[J]. Chinese Science Bulletin, 56(1): 64-69. doi: 10.1007/s11434-010-4235-8 [13] LI Y W, XU Q H, ZHANG S R, et al. , 2019. Significance of pollen assemblages for the vegetationcomposition of alpine shrub meadow in the Qinghai-Tibetan Plateau, China[J]. Chinese Science Bulletin, 64(20): 2141-2150. (in Chinese with English abstract) doi: 10.1360/N972019-00126 [14] LI Z H, JIANG B Y, DONG X P, et al. , 2020. Collapses of loess at the front of the Emei tableland in the Yuncheng basin and their major geological controlling factors[J]. Coal Geology & Exploration, 48(2): 171-178. (in Chinese with English abstract) [15] LIN X D, YUAN H Y, XU P, et al. , 2017. Zonational characteristics of earthquake focal mechanism solutions in North China[J]. Chinese Journal of Geophysics, 60(12): 4589-4622. (in Chinese with English abstract) [16] LIU Z S, 2000. Early cretaceous sporopollen assemblage from the Hanxia of Yumen in Gansu, NW China[J]. Acta Micropalaeontologica Sinica, 17(3): 73-84. (in Chinese with English abstract) [17] LUO C X, PAN A D, ZHENG Z, 2006. Progresses about the studies on the relationship between topsoil spore-pollen and vegetation in arid areas of northwest China[J]. Arid Zone Research, 23(2): 314-319. (in Chinese with English abstract) [18] MINCKLEY T, WHITLOCK C, 2000. Spatial variation of modern pollen in Oregon and southern Washington, USA[J]. Review of Palaeobotany and Palynology, 112(1-3): 97-123. doi: 10.1016/S0034-6667(00)00037-3 [19] MOLNAR P, 2005. Mio-Pliocene growth of the Tibetan Plateau and evolution of East Asian climate[J]. Palaeontologia Electronica, 8(1): 1-23. [20] National Stratigraphic Committee, 1963. Cenozoic erathem of China[M]. Beijing: Science Press. (in Chinese) [21] Palynological Section, Morphological Department, Institute of Botany, Chinese Academy of Sciences, 1960. Pollen morphology of Chinese plants[M]. Beijing: Science Press. (in Chinese) [22] POKROVSKAYA, GESHIDEKOVA, GRICHUK, et al. , 1956. Pollen analysis[M]. WANG F X, ZHANG J T, MA Y Q, et al, trans. Beijing: Science Press. (in Chinese) [23] QI Y, XU H B, ZHANG J X, et al. , 2011. Geochemistry, geochronology and geological significance of Gufengshan granodiorite in Linfen Grabben Basin[J]. Geological Review, 57(4): 565-573. (in Chinese with English abstract) [24] QI Y, LUO J H, WU J D, et al. , 2016. Geochemical and Sr-Nd-Pb isotopic composition of the Canfang and Gufengshan granodiorite plutons in central-southern North China[J]. Acta Petrologica Sinica, 32(7): 2015-2028. (in Chinese with English abstract) [25] QIN B C, FANG W X, ZHANG J G, et al. , 2021. Quaternary sedimentary sequence and sedimentary environment restoration in the Jinzhong Basin, Fenhe Rift Valley[J]. Journal of Geomechanics, 27 (6): 1035-1050. (in Chinese with English abstract) [26] QIU D W, GONG W B, YAN J Y, et al. , 2021. Geological environment changes during the late Pleistocene-Holocene on the E'mei tableland in the northern Yuncheng basin, Shanxi Province: implications for the distribution of human settlements[J]. Journal of Geomechanics, 27(2): 326-338. (in Chinese with English abstract) [27] SCHÄBITZ F, 1994. Holocene climatic variations in northern Patagonia, Argentina[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 109(2-4): 287-294. doi: 10.1016/0031-0182(94)90180-5 [28] Shanxi Provincial Geological Bureau 214 Geological Team, 1982. Instructions for 1: 100000 Geological Map of Yuncheng District, Shanxi Province[M]. Yuncheng: Water Resources Bureau of Yuncheng District Administration. (in Chinese) [29] Shanxi Provincial Geological Prospecting Bureau, 1989. Regional geology of Shanxi province[M]. Beijing: Geology Press. (in Chinese) [30] SHI W, DONG S W, HU J M, 2020. Neotectonics around the Ordos Block, North China: a review and new insights[J]. Earth-Science Reviews, 200: 102969. doi: 10.1016/j.earscirev.2019.102969 [31] SONG Z C, 1999. Fossil spores and pollen of China (Vol. 1)[M]. Beijing: Science Press. (in Chinese) [32] Sporopollen Group, Paleobotany Research Office, Beijing Institute of Botany, Chinese Academy of Sciences, 1976. Spore morphology of ferns in China[M]. Beijing: Science Press. (in Chinese) [33] SU P, HE H L, TAN X B, et al, 2021. Initiation and evolution of the Shanxi rift system in North China: evidence from low-temperature thermochronology in a plate reconstruction framework[J]. Tectonics, 40(3): e2020TC006298. [34] SUO Y H, LI S Z, CAO X Z, et al. , 2017. Mesozoic-Cenozoic inversion tectonics of East China and its implications for the subduction process of the oceanic plate[J]. Earth Science Frontiers, 24(4): 249-267. (in Chinese with English abstract) [35] TONG Y S, ZHENG S H, QIU Z D, 1995. Cenozoic mammals ages of China[J]. Vertebrata Palasiatica, 33(4): 290-314. (in Chinese with English abstract) [36] XU Q H, LI Y C, YANG X L, et al. , 2005. A study of some typical pollen types taphonomy and relationships with vegetation in the northeast of Qinghai-Tibet Plateau[J]. Advances in Earth Science, 20(1): 89-98. (in Chinese with English abstract) [37] XU Q H, LI Y C, YANG X L, et al. , 2007. Quantitative relationship between pollen and vegetation in northern China[J]. Science in China Series D: Earth Sciences, 50(4): 582-599. doi: 10.1007/s11430-007-2044-y [38] XU Z L, 2015. Oligocene-Miocene pollen records in Xunhua Basin, northeastern Tibetan Plateau and its implications for evolution of the East Asian monsoon[D]. Wuhan: China University of Geosciences. (in Chinese with English abstract) [39] YAN J Y, 2021. Late Cenozoic tectonic-sedimenatary, uplifting and denudational process of the Yuncheng Basin and northern Gushan Mountain[D]. Beijing: Chinese Academy of Geological Sciences. (in Chinese with English abstract) [40] YAN S, 1991. The characteristics of Quaternary sporo-pollen assemblage and the vegetation succesion in Xinjiang[J]. Arid Land Geography, 14(2): 1-9. (in Chinese with English abstract) [41] ZDANSKY O, 1923. Fundorte der Hipparion-Fauna um Pao-Te-Hsien in NW-Shansi[J]. Bulletin of the Geological Survey, China, 5: 69-81. [42] ZHANG Y X, YUE L P, CHEN D L, et al. , 1997. Petrographic marks for stratigraphic division of the Neogene Red Clay in North China and their significance[J]. Journal of Stratigraphy, 21(1): 63-67. (in Chinese with English abstract) [43] ZHONG Q M, SHAO B, HOU G T, 2022. Numerical simulation and analysis of lithospheric stress field in Fenwei graben[J]. Progress in Geophysics, 37(1): 152-163. (in Chinese with English abstract) [44] 陈兴强, 2017. 汾渭地堑系新生代沉积-构造演化及其形成机制[D].博士学位论文.北京: 北京大学. [45] 邓涛, 王伟铭, 岳乐平, 等, 2004. 新近系保德阶建阶研究新进展[J]. 地层学杂志, 28(1): 41-47. [46] 邓涛, 王伟铭, 岳乐平, 2008. 中国陆相新近系山旺阶和保德阶综合研究报告[M]. 见: 第三届全国地层委员会编. 中国主要断代地层建阶研究报告(2001~2005). 北京: 地质出版社, 13-31. [47] 方小敏, 2017. 青藏高原隆升阶段性[J]. 科技导报, 35(6): 42-50. [48] 韩晓明, 刘芳, 张文韬, 等, 2015. 基于震源机制资料分析河套地震带的应力场变化特征[J]. 地震地质, 37(4): 1030-1042. [49] 寇琳琳, 李振宏, 董晓朋, 等, 2021. 青藏高原东北缘隆德观音店剖面碎屑锆石年龄序列及地质意义[J]. 地质力学学报, 27(6): 1051-1064. [50] 李三忠, 曹现志, 王光增, 等, 2019. 太平洋板块中-新生代构造演化及板块重建[J]. 地质力学学报, 25(5): 642-677. [51] 李文漪, 1998. 中国第四纪植被与环境[M]. 北京: 科学出版社. [52] 李怡雯, 徐海清, 张生瑞, 等, 2019. 青藏高原高寒灌丛草甸花粉组合的植被指示性[J]. 科学通报, 64: 2141-2150. [53] 李振宏, 姜博宇, 董晓朋, 等, 2020. 运城盆地峨眉台地前缘黄土塌陷现状及地质主控因素[J]. 煤田地质与勘探, 48(2): 171-178. [54] 林向东, 袁怀玉, 徐平, 等, 2017. 华北地区地震震源机制分区特征[J]. 地球物理学报, 60(12): 4589-4622. [55] 刘兆生, 2000. 甘肃玉门旱峡早白垩世孢粉组合[J]. 微体古生物学报, 17(3): 73-84. [56] 罗传秀, 潘安定, 郑卓, 2006. 西北干旱地区表土孢粉与植被关系研究进展[J]. 干旱区研究, 23(2): 314-319. [57] 坡克罗夫斯卡娅, 格刺德科娃, 格里丘克, 等, 1956. 花粉分析[M]. 王伏雄, 张金谈, 马毓泉, 等译. 北京: 科学出版社. [58] 齐玥, 徐鸿博, 张竞雄, 等, 2011. 临汾断陷盆地孤峰山花岗闪长岩的地球化学和年代学及其地质意义[J]. 地质论评, 57(4): 565-573. [59] 齐玥, 罗金海, 巫嘉德, 等, 2016. 华北中南部蚕坊和孤峰山花岗闪长岩体的地球化学特征和Sr-Nd-Pb同位素组成[J]. 岩石学报, 32(7): 2015-2028. [60] 秦帮策, 方维萱, 张建国, 等, 2021. 汾河裂谷晋中盆地内第四纪沉积序列与沉积环境恢复[J]. 地质力学学报, 27 (6): 1035-1050. [61] 仇度伟, 公王斌, 闫纪元, 等, 2021. 山西运城盆地北部峨嵋台地晚更新世: 全新世地质环境变化及其对人类聚落分布的影响[J]. 地质力学学报, 27(2): 326-338. [62] 全国地层委员会, 1963. 全国地层会议学术报告汇编: 中国的新生界[M]. 北京: 科学出版社. [63] 山西省地质局二一四地质队, 1982. 山西省运城地区地质图系说明书[M]. 运城: 运城地区行署水利局. [64] 山西省地质矿产局, 1989. 山西省区域地质志[M]. 北京: 地质出版社. [65] 宋之琛, 1999. 中国孢粉化石(第一卷): 晚白垩世和第三纪孢粉[M]. 北京: 科学出版社. [66] 索艳慧, 李三忠, 曹现志, 等, 2017. 中国东部中新生代反转构造及其记录的大洋板块俯冲过程[J]. 地学前缘, 24(4): 249-267. [67] 童永生, 郑绍华, 邱铸鼎, 1995. 中国新生代哺乳动物分期[J]. 古脊椎动物学报, 33(4): 290-314. [68] 许清海, 李月丛, 阳小兰, 等, 2005. 青藏高原东北部典型花粉类型埋藏特征及其与植被关系的研究[J]. 地球科学进展, 20(1): 89-98. [69] 徐增连, 2015. 青藏高原东北缘循化盆地渐新世-中新世孢粉记录与东亚季风演化[D]. 武汉: 中国地质大学. [70] 闫纪元, 2021. 运城盆地及北侧孤山晚新生代构造-沉积与隆升-剥蚀过程研究[D]. 北京: 中国地质科学院. [71] 闫顺, 1991. 新疆第四纪孢粉组合特征及植被演替[J]. 干旱区地理, 14(2): 1-9. [72] 张云翔, 岳乐平, 陈丹玲, 等, 1997. 中国北部新第三纪红层划分的岩石学标志及其意义[J]. 地层学杂志, 21(1): 63-67. [73] 中国科学院北京植物研究所古植物研究室孢粉组, 1976. 中国蕨类植物孢子形态[M]. 北京: 科学出版社. [74] 中国科学院植物研究所形态室孢粉组, 1960. 中国植物花粉形态[M]. 北京: 科学出版社. [75] 中国植被编辑委员会, 1980. 中国植被[M]. 北京: 科学出版社. [76] 仲启蒙, 邵博, 侯贵廷, 2022. 汾渭地堑岩石圈的应力场数值模拟与分析[J]. 地球物理学进展, 37(1): 152-163. -
 下载:
下载:


计量
- 文章访问数: 1410
- HTML全文浏览量: 545
- PDF下载量: 103
- 被引次数: 0
